



IMPROVE DRY MATTER INTAKE
Nutritionists and dairymen alike have been trying to maximize dry matter intake of their lactating cow diet.
Allen, 2000 stated, “Energy intake is a primary limitation on milk yield for high producing dairy cows and is determined by net energy content of the diet and DMI. The maximal productive capacity of an animal will depend on its genetic potential and will vary over the animal’s lifetime according to its age, physiological status (e.g. lactating, pregnant), and climate. Each animal has a maximal rate at which it can utilize nutrients and metabolic fuels and unless DMI is limited by physical capacity, mechanisms must exist that balance supply with demand for nutrients.” Allen, 2000
Dry matter intake (DMI) is determined by meal size and meal frequency that are influenced by animal and dietary factors affecting satiety and hunger. Certain feed additives and nutrients can cause changes in DMI. The interest of this publication is to explore fatty acids (FA) and their effect on DMI.
Allen, 2000 published a review on factors affecting feed intake including certain FA. Supplemental fat sometimes depresses feed intake due to effects of fat on ruminal fermentation and gut motility, acceptability of diets containing added fat, release of gut hormones, and oxidation of fat in the liver. Significant decreases in DMI were observed for Ca-PFA for 11 of 24 comparisons. In addition, Ca-PFA resulted in a numerical decrease in DMI in 22 of the 24 comparisons. Other sources of fat had less consistent effects on DMI and there were few significant decreases in DMI by added FA for each category of fat. A summary of these findings is illustrated in Table 1.

Table 1. Effects of fatty acid source on DMI in Lactating cows. (adapted from Allen, 2000).
The effects of Ca salts of PFAD (CAS) on DMI reduction was linear indicating significance when included as low as 1% of the diet DM. The data shows us that fat alone is not responsible for the reduction in DMI. The composition of FA within in a fat source appears to be the cause. Allen 2000 concluded “ Although energy utilization is more efficient for digested fat than digested carbohydrate, it is clear that addition of fat to the diet does not always result in increased net energy intake and that reduction in DMI is one of the primary reasons.
There have been several meta-analyses regarding the effects FA on DMI depression. These results are shown in Table 2.
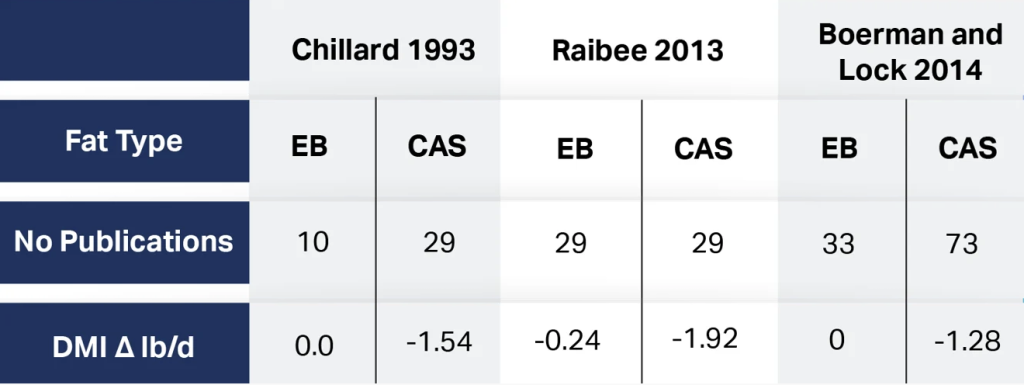
These meta-analyses all indicate that Ca salts of PFAD (CAS) cause significant reduction in DMI based on dozens of published reports. The question then becomes what physical or chemical characteristics lead to the issues with DMI depression. Palatability of fat sources has been researched by Grummer et al. 1990 as a possible factor involved in decreased DMI from fat sources. They reported that some fat sources tested reduced DMI initially, but cows became adjusted after a period of time except for one—CAS. Cows do not become accustomed to CAS presence in the diet. Some other factor is causing DMI depression with respect to CAS.
Degree of unsaturation of FA has been another potential factor involved in FA effect on DMI. DMI depression has been observed with increased degree of unsaturation. Drackley et al 1992 observed when unsaturated LCFA were infused into the abomasum, increased degree of unsaturation reduced DMI more than saturated FA. These researchers suggested that unsaturated LCFA reaching the small intestine of dairy cows affects gastrointestinal motility and DMI. That brings us to another potential mechanism involving gut hormones such as CCK and GLP-1.
Choi and Palmquist 1996 observed increases in CCK and a consequent reduction in DMI when 0, 3%, 6%, or 9% CAS was added to the diet. Fat supplementation increased post-feeding plasma cholecystokinin concentrations and linearly increased plasma pancreatic polypeptide (GLP-1) concentrations. The effects of CAS on CCK plasma concentrations are reported in Figure 1.
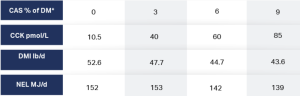

There are at least 10 hormones that activate the stomach and intestines, induce gastric secretions, regulate glucose metabolism, and influence satiety centers in the brain (Song et al 2015). Among these CCK and GLP-1 are the primary hormones affecting satiety. Administration of CCK to humans and animals reduces meal size and duration. CCK causes sensations of satiety as well as reducing gastric emptying and stimulating pancreatic enzyme secretions (Wren and Bloom 2007). When specific FA, such as palmitic and oleic acids, empty into the duodenum, CCK receptors begin sending satiety signals to the brain reducing gastric emptying which, in turn, reduces meal size and duration. This happens every time these FA enter the duodenum in sufficient quantity. The sequence of events is controlled and modulated hormonally.
Published research work by Relling and Reynolds 2007 and Bradford et al 2008 in lactating dairy cows has shown similar results comparing EB 100 to CAS. EB 100 is a FA source containing mostly saturated FA high in stearic acid while CAS is primarily palmitic and oleic acids. CAS significantly increased CCK while EB 100 was similar to the control. Piantoni et al 2013 observed a significant increase in CCK comparing an 85% palmitic acid FA source to the control. These data suggest that FA sources high in palmitic and oleic acids increase CCK potentially leading to DMI depression, while a FA source high in stearic acid does not. These reports are shown in Table 3.

These published reports show the mechanism by which specific FA such as palmitic and oleic cause increases in CCK when fed to lactating cows. This in turn may lead to depressed DMI. To further investigate this hormonally modulated effect, a meta-analysis performed by Sellers et al. 2017 compared FA sources to the control. These findings are in Table 4.
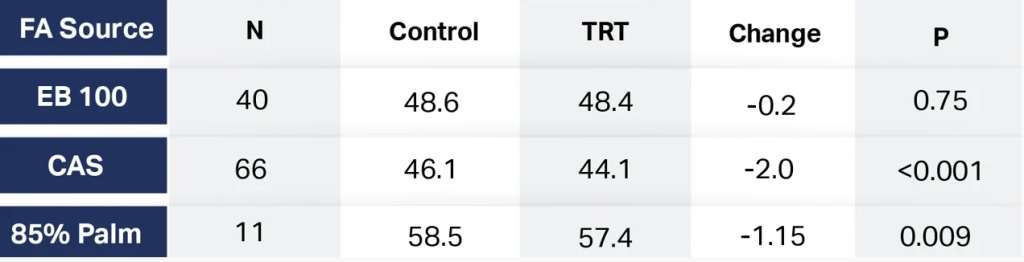
The advantage that EB 100 has over CAS and 85% Palm is improved DMI. This has multiple positive effects ranging from higher NEL intake, higher milk yield, better BCS and body weight gain throughout lactation, improved reproductive parameters from conception rates to days open, and higher value as measured by IOFC. This is a mammalian phenomenon. The appetite of humans, mice, cows, apes, pigs, etc. are all affected similarly by these specific FA. EB 100 is the consultant’s choice for improved performance from milking to breeding. Contact your Actus Nutrition representative for more details on their Energy Booster line of products.
References available upon request.